
Flagging the HIV reservoir: a potential new marker for CD4 cells hiding HIV
24 April 2017. Related: Cure-related research, Basic science and immunology.
Richard Jefferys, TAG
The rarity of CD4 T cells containing latent HIV in people on antiretroviral therapy – the typical estimate is around one per million CD4 T cells – makes them extremely challenging both to study and to target with therapies.
A paper published online yesterday in Nature represents a possible breakthrough in this area, reporting that it may be possible to identify many latently infected CD4 T cells due to expression of a particular cell surface protein, CD32a. [1]
The paper’s authors, led by Benjamin Descours from the laboratory of Monsef Benkirane at Université de Montpellier, made their discovery using an in vitro model they developed that allows for the direct infection of resting CD4 T cells by a modified version of HIV. Descours and colleagues used the system to generate latently infected CD4 T cells and then looked at whether any genes in these cells were behaving differently compared to uninfected CD4 T cells.
Out of 103 genes upregulated exclusively in the infected cells, ١٦ were selected for further study because they encode cell surface proteins that can be used to rapidly sort cells using a flow cytometer. The gene FCGR2A turned out to be most strongly and consistently upregulated, which encodes the cell surface receptor CD32a.
The researchers found that when resting CD4 T cells sampled from uninfected donors were latently infected with HIV in the laboratory, the expression of CD32a was reliably induced: approximately 90% of the CD32a+ CD4 T cells generated in these experiments contained latent HIV. Furthermore, treatment of the samples with the integrase inhibitor raltegravir before infection prevented CD32a expression, suggesting that the integration of HIV into the CD4 T cell genome was causing the receptor to be expressed.
To try and confirm the relevance of the laboratory findings, CD4 T cells from 12 HIV positive individuals on suppressive ART were sampled and sorted based on CD32a expression. When the amount of HIV DNA was compared between subsets, there was a significant concentration of latent HIV infection in CD4 T cells with the highest levels of CD32a expression (an approximately 1,024-fold enrichment of HIV DNA in CD4 T cells with high CD32a expression versus those lacking CD32a).
There was variation between participants, however, with the contribution of the CD32a+ CD4 T cell population to the total HIV DNA reservoir ranging from 26.8% to 86.3% – the average contribution was a little over 50%. A similar concentration of the HIV reservoir in CD32a+ CD4 T cells was also documented with an assay measuring replication-competent HIV rather than HIV DNA.
The researchers highlight several potentially important implications of these findings:
- Sorting CD4 T cells based on CD32a expression should offer an easier means of studying the HIV reservoir than has previously been available, facilitating studies at the single-cell level.
- The normal biological function of CD32a involves recognising antigen-antibody complexes via the Fc region of antibodies and delivering signals capable of activating a broad spectrum of immune responses. This suggests CD32a might have a role in mediating responses to broadly neutralising anti-HIV antibodies, (bNAbs) and the potential to contribute to clearance of reservoir cells by these antibodies (evidence of bNAbs contributing to HIV reservoir depletion has been reported in a mouse model. [2]
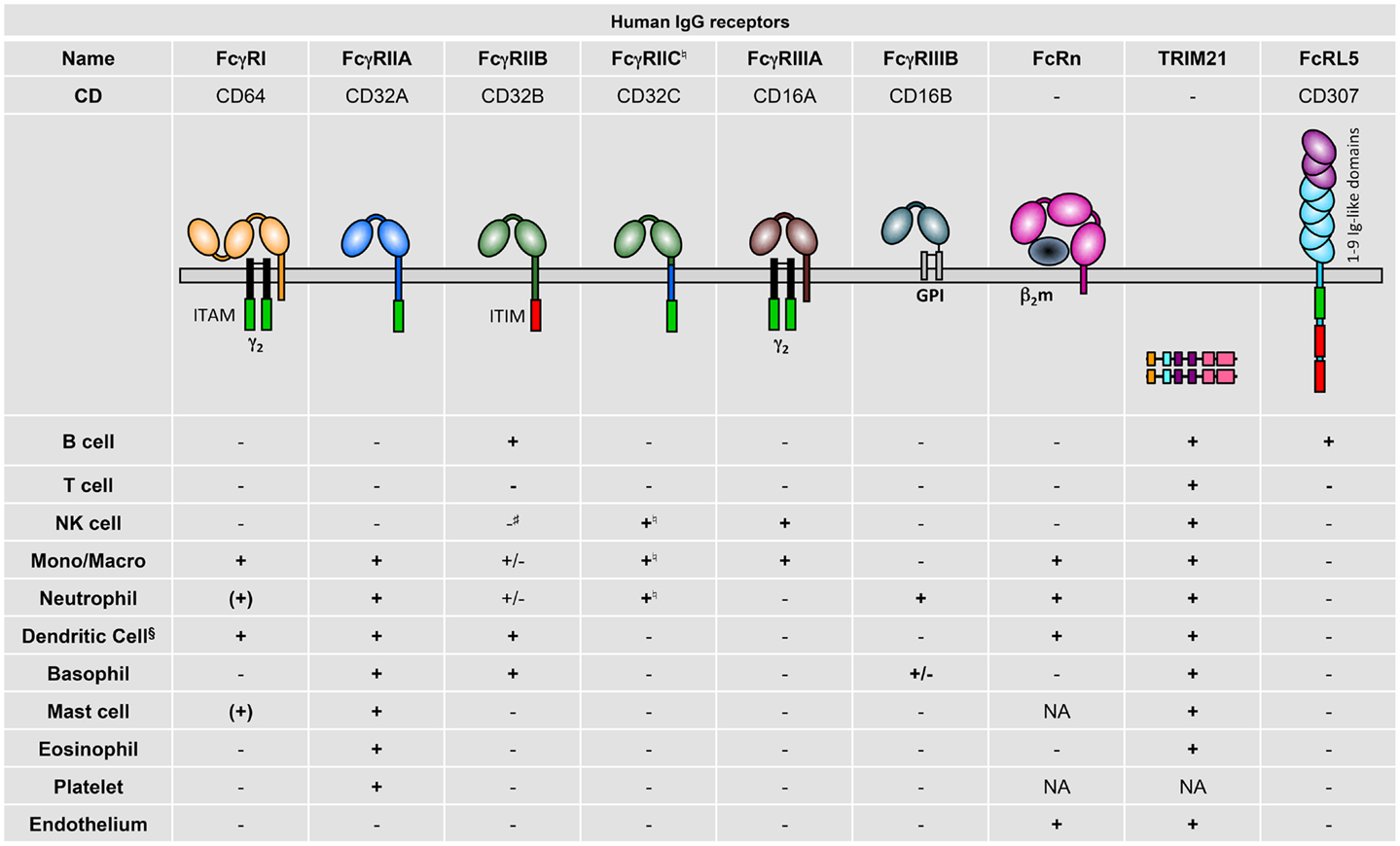
- CD32a may allow for direct targeting of a large portion of the HIV reservoir in CD4 T cells with elimination strategies. However, as Doug Richman notes in an accompanying commentary in Nature [3], CD32a expression on other cell types (see figure 2 of this 2014 review [4, 5]) raises concerns as to whether this approach could be pursued safely.
The study also raises some technical questions about the mechanism by which HIV latency is established. Under most circumstances, CD4 T cells need to be activated to be susceptible to HIV infection, and viral latency has generally been thought to result from some infected cells returning to a resting state with HIV integrated into their genomes. [6]
But the laboratory model used by Descours et al involves manipulations that allow direct infection of resting CD4 T cells, and the researchers did not see CD32a expression in CD4 T cells that were infected after activation. This poses the question of whether the latently infected CD32a-expressing CD4 T cells sampled from people on ART were infected while they were in a resting state, or if CD32a can eventually be expressed when an HIV-infected, activated CD4 T cell returns to a resting state. Additional investigations will be required to address this issue.
Source:
TAG Basic Science Blog (16 March 2017)
http://tagbasicscienceproject.typepad.com
References:
- Descours B et al. CD32a is a marker of a CD4 T-cell HIV reservoir harbouring replication-competent proviruses. Nature (2017) doi:10.1038/nature21710
http://www.nature.com/nature/journal/vaop/ncurrent/full/nature21710.html - Horwitz J A et al. HIV-1 suppression and durable control by combining single broadly neutralizing antibodies and antiretroviral drugs in humanized mice. PNAS (October 2013): 110(41);16538-16543. doi: 10.1073/pnas.1315295110
http://www.pnas.org/content/110/41/16538.abstract - Richman D.HIV: Finding latent needles in a haystack. Nature (2017) doi:10.1038/nature21899
http://www.nature.com/nature/journal/vaop/ncurrent/full/nature21899.html - Gillis C et al. Contribution of human FcγRs to disease with evidence from human polymorphisms and transgenic animal studies. Front Immunol (May 2014). doi.org/10.3389/fimmu.2014.00254.
http://journal.frontiersin.org/article/10.3389/fimmu.2014.00254/full - Ibid, Gillis et al. Figure 2. Human IgG receptor expression pattern.
http://www.frontiersin.org/files/Articles/89310/fimmu-05-00254-HTML/image_m/fimmu-05-00254-g002.jpg (JPG) - Persoud D et al. Latency in Human Immunodeficiency Virus type 1 infection: no easy answers. J. Virol. (February 2003) 77(3);1659-1665. doi: 10.1128/JVI.77.3.1659-1665.2003
http://jvi.asm.org/content/77/3/1659
{kind=link}